Mechanism of action (MoA) animations for selected surface receptors in the 3D real-time interactive virtual cell environment Nanoscape
Protein structures were retrieved from the RSCB protein data bank (PDB) and MoA
animations based on data from scientific papers were simulated using the mMaya modelling and rigging kits (Clarafi). Rigged surface or backbone meshes were extracted, animation playblasts recorded in Maya (Autodesk) and composited in After Effects (Adobe CC).
animations based on data from scientific papers were simulated using the mMaya modelling and rigging kits (Clarafi). Rigged surface or backbone meshes were extracted, animation playblasts recorded in Maya (Autodesk) and composited in After Effects (Adobe CC).
Created by Shereen Kadir, 3DVAL, UNSW Art and Design, Sydney, Australia, 2019-20
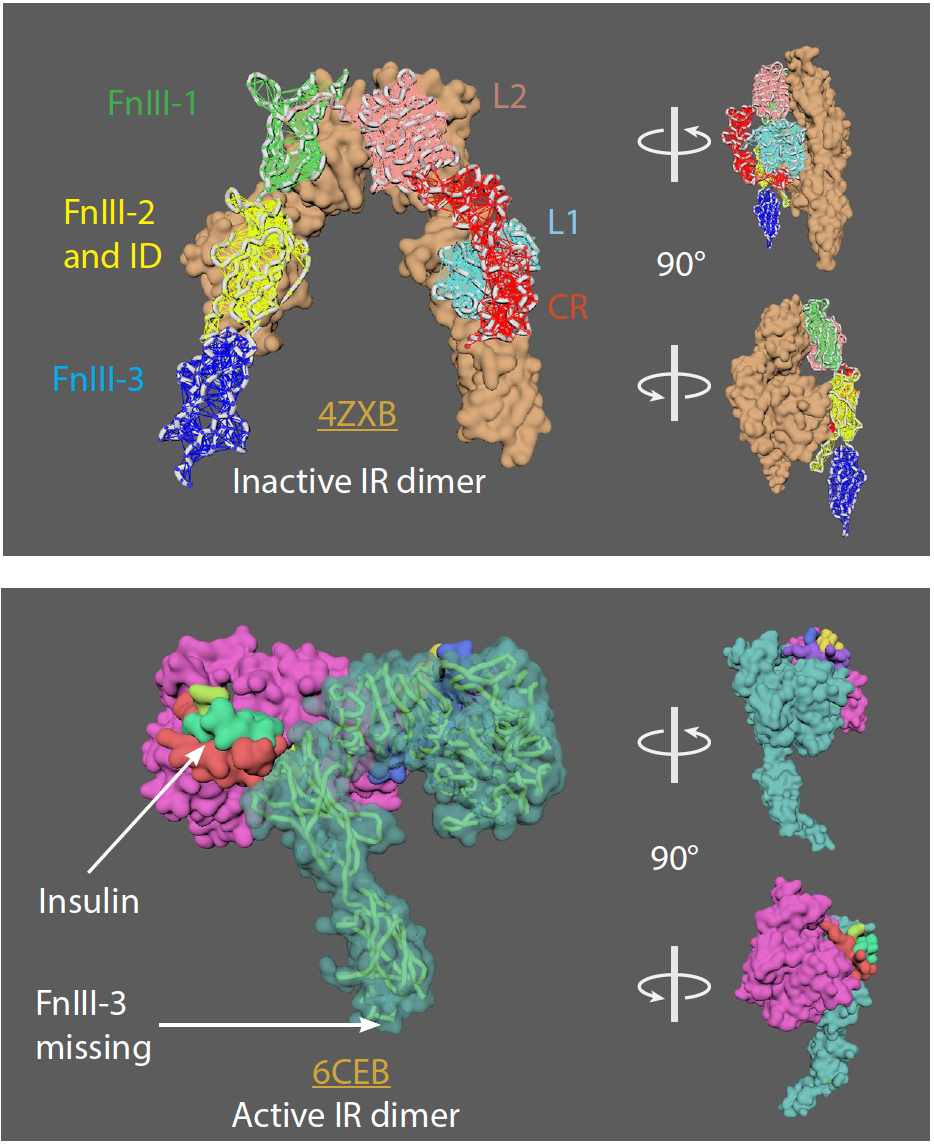
Insulin Receptor (IR)
• IR is a dimer of heterodimers that comprises two α-chains and two β-chains, represented as (αβ)2.
• 2 insulin molecules per dimer. Each bound between L1 of one monomer and FnIII-1 of the other momoner.
• IR goes from an inverted U-shaped dimer to a T-shaped dimer upon ligand binding. One ligand binding to one receptor can lead to the conformational change.
• 2 insulin molecules per dimer. Each bound between L1 of one monomer and FnIII-1 of the other momoner.
• IR goes from an inverted U-shaped dimer to a T-shaped dimer upon ligand binding. One ligand binding to one receptor can lead to the conformational change.
References:
1. Croll, T., Smith, B., Margetts, M. et al. Higher-Resolution Structure of the Human Insulin Receptor Ectodomain: Multi-Modal Inclusion of the Insert Domain. Structure, 24 (3), 469-476. (2016) https://doi.org/10.1016/j.str.2015.12.014
2. Scapin, G., Dandey, V., Zhang, Z. et al. Structure of the insulin receptor–insulin complex by single-particle cryo-EM analysis. Nature 556, 122–125 (2018). https://doi.org/10.1038/nature26153
3. Gutmann, T., Kim, K., Grzybek, M. et al. Visualization of ligand-induced transmembrane signaling in the full-length human
insulin receptor. J Cell Biol 7; 217 (5): 1643–1649 (2018) https://doi.org/10.1083/jcb.201711047
1. Croll, T., Smith, B., Margetts, M. et al. Higher-Resolution Structure of the Human Insulin Receptor Ectodomain: Multi-Modal Inclusion of the Insert Domain. Structure, 24 (3), 469-476. (2016) https://doi.org/10.1016/j.str.2015.12.014
2. Scapin, G., Dandey, V., Zhang, Z. et al. Structure of the insulin receptor–insulin complex by single-particle cryo-EM analysis. Nature 556, 122–125 (2018). https://doi.org/10.1038/nature26153
3. Gutmann, T., Kim, K., Grzybek, M. et al. Visualization of ligand-induced transmembrane signaling in the full-length human
insulin receptor. J Cell Biol 7; 217 (5): 1643–1649 (2018) https://doi.org/10.1083/jcb.201711047
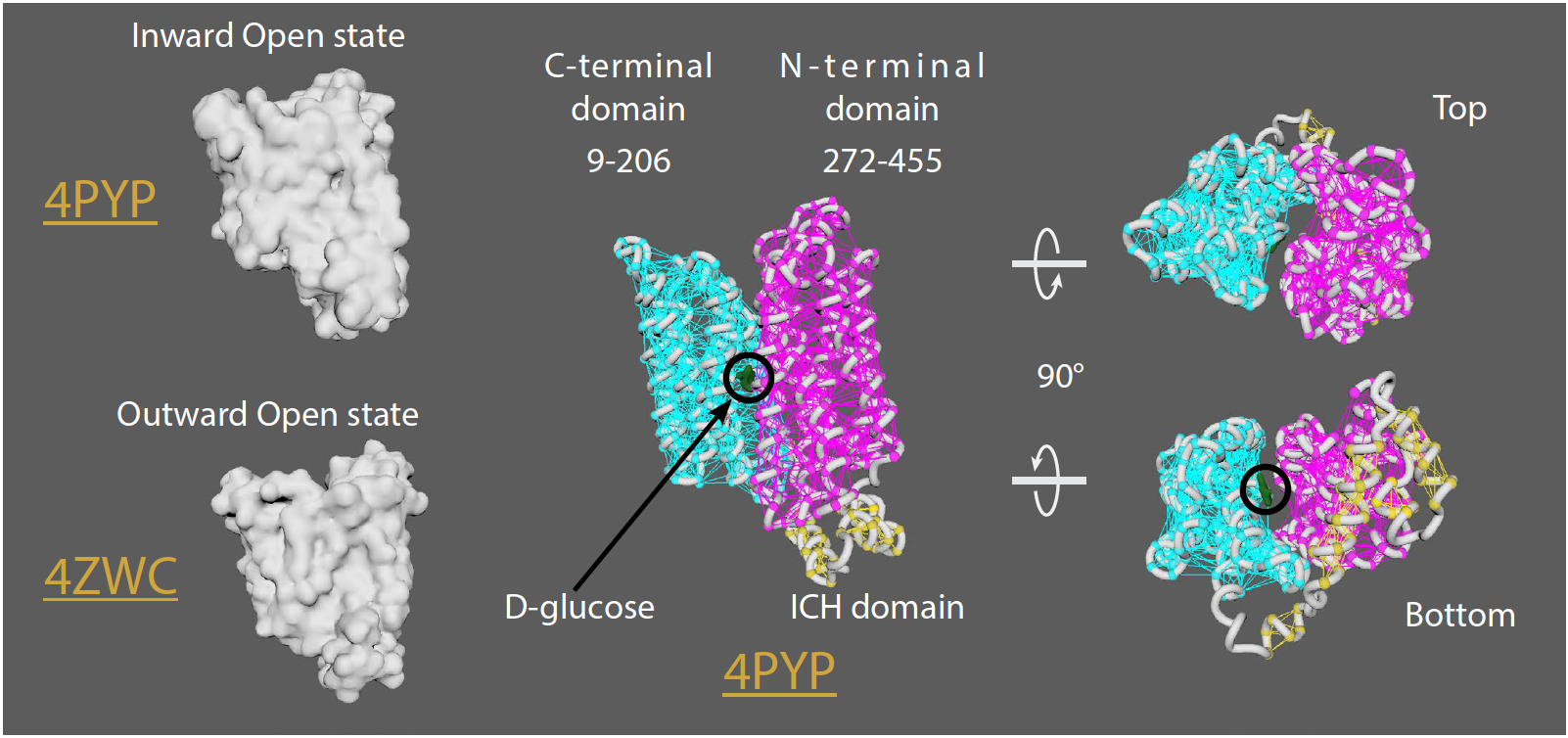
GLUT1 (glucose transporter 1)
• GLUT1 is over-expressed in many cancer cells
• 12 transmembrane (TM) segments form the N-terminal and C-terminal domains
• Rotation of the N-terminal and C-terminal domains occurs to transition from an outwardopen
to an outward-occluded, and finally an inward-open state to allow D-glucose transport
• 12 transmembrane (TM) segments form the N-terminal and C-terminal domains
• Rotation of the N-terminal and C-terminal domains occurs to transition from an outwardopen
to an outward-occluded, and finally an inward-open state to allow D-glucose transport
References:
1. Deng, D., Sun, P., Yan, C. et al. Molecular basis of ligand recognition and transport by glucose transporters. Nature
526, 391–396 (2015). https://doi.org/10.1038/nature14655
2. Deng, D., Xu, C., Sun, P. et al. Crystal structure of the human glucose transporter GLUT1. Nature 510, 121–125
(2014). https://doi.org/10.1038/nature13306
1. Deng, D., Sun, P., Yan, C. et al. Molecular basis of ligand recognition and transport by glucose transporters. Nature
526, 391–396 (2015). https://doi.org/10.1038/nature14655
2. Deng, D., Xu, C., Sun, P. et al. Crystal structure of the human glucose transporter GLUT1. Nature 510, 121–125
(2014). https://doi.org/10.1038/nature13306
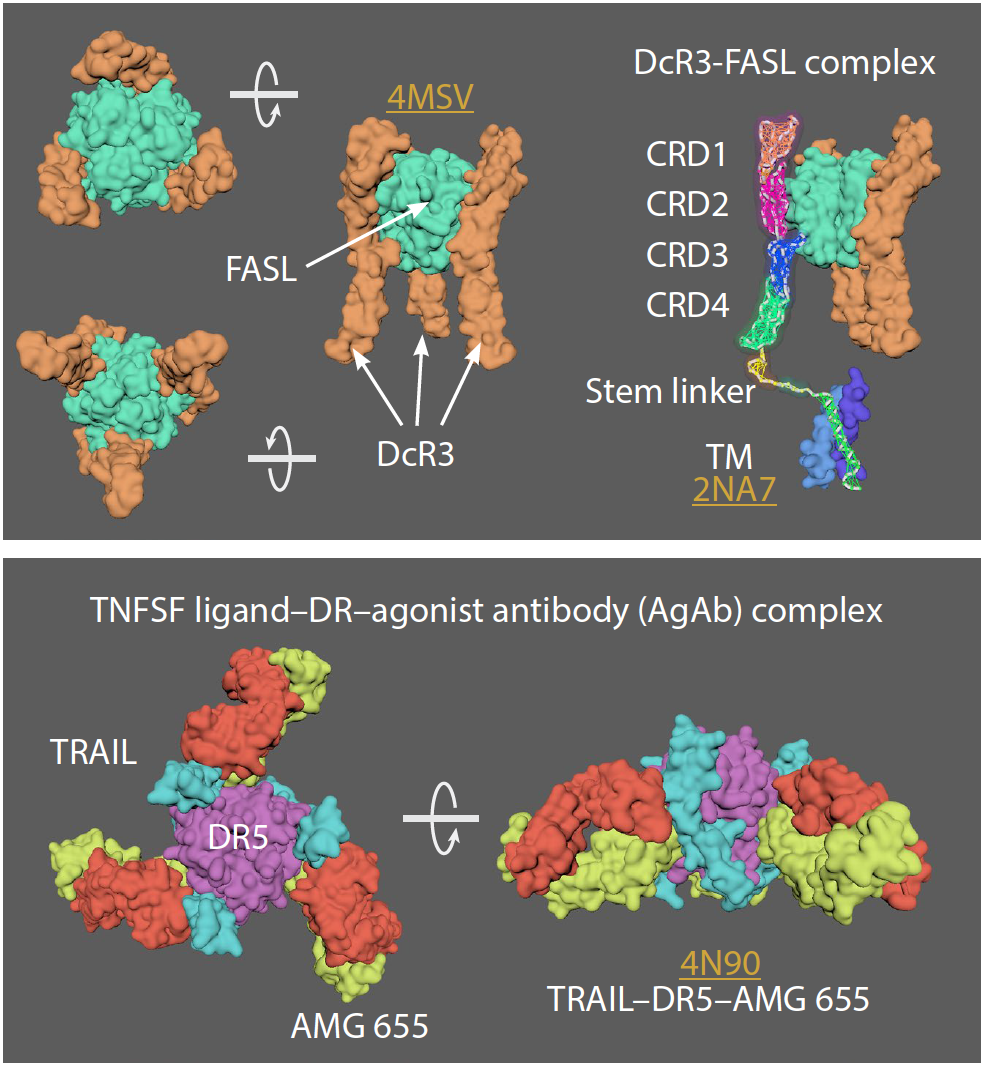
TNFR super family
• In the TNFR superfamily there are 3 groups of receptors: 1) Death receptors (DRs); 2) TNFR-associated factor (TRAF)–interacting receptors, and 3) decoy receptors (DcRs)
• Basic unit of signalling is a trimeric ligand and three receptors.
• Each receptor binds on the outside interface of 2 ligand monomers
• Receptors can pre-assemble (as resting or “nonsignaling” state) on the cell surface and can form either parallel or antiparallel dimers.
• Preferred model is antiparallel dimers
• Antiparallel dimers are formed between the receptor CRD1 and CRD2 domains which occludes the ligand-binding site.
• Arranged as a large hexagonal lattice where each point connects 3 receptor monomers.
• Individual ligand-receptor complexes ~120 Å apart. Total edge length of ~170 Å (may vary with receptor type).
• Ligand binding leads to a conformational change in receptors (into an upright position, perpendicular to the cell surface) allowing cell signaling but maintaining hexagonal symmetry.
• Basic unit of signalling is a trimeric ligand and three receptors.
• Each receptor binds on the outside interface of 2 ligand monomers
• Receptors can pre-assemble (as resting or “nonsignaling” state) on the cell surface and can form either parallel or antiparallel dimers.
• Preferred model is antiparallel dimers
• Antiparallel dimers are formed between the receptor CRD1 and CRD2 domains which occludes the ligand-binding site.
• Arranged as a large hexagonal lattice where each point connects 3 receptor monomers.
• Individual ligand-receptor complexes ~120 Å apart. Total edge length of ~170 Å (may vary with receptor type).
• Ligand binding leads to a conformational change in receptors (into an upright position, perpendicular to the cell surface) allowing cell signaling but maintaining hexagonal symmetry.
References:
1. Liu, W., Ramagopal, U., Cheng, H. et al. Crystal Structure of the Complex of Human FasL and Its Decoy Receptor DcR3. Structure, 24 (11), 2016-2023, (2016). https://doi.org/10.1016/j.str.2016.09.009
2. Fu, Q., Fu, T., Cruz, A. et al. Structural Basis and Functional Role of Intramembrane Trimerization of the Fas/CD95 Death Receptor. Molecular Cell, 61 (4), 602-613, (2016). https://doi.org/10.1016/j.molcel.2016.01.009
3. Graves, J., Kordich, J., Huang, T. et al. Apo2L/TRAIL and the Death Receptor 5 Agonist Antibody AMG 655 Cooperate to Promote Receptor Clustering and Antitumor Activity. Cancer Cell, 26 (2), 177-189, (2014). https://doi.org/10.10/j.ccr.2014.04.028
4. Naismith, J., Devine, T., Kohno, T. et al. Structures of the extracellular domain of the type I tumor necrosis factor receptor. Structure, 4 (11), 1251-1262, (1996). https://doi.org/10.1016/S0969-2126(96)00134-7
5. Vanamee, É. and Faustman, D. Structural principles of tumor necrosis factor superfamily signaling. Science Signaling, 11 (511), eaao4910, (2018) https://doi.org/10.1126/scisignal.aao4910
1. Liu, W., Ramagopal, U., Cheng, H. et al. Crystal Structure of the Complex of Human FasL and Its Decoy Receptor DcR3. Structure, 24 (11), 2016-2023, (2016). https://doi.org/10.1016/j.str.2016.09.009
2. Fu, Q., Fu, T., Cruz, A. et al. Structural Basis and Functional Role of Intramembrane Trimerization of the Fas/CD95 Death Receptor. Molecular Cell, 61 (4), 602-613, (2016). https://doi.org/10.1016/j.molcel.2016.01.009
3. Graves, J., Kordich, J., Huang, T. et al. Apo2L/TRAIL and the Death Receptor 5 Agonist Antibody AMG 655 Cooperate to Promote Receptor Clustering and Antitumor Activity. Cancer Cell, 26 (2), 177-189, (2014). https://doi.org/10.10/j.ccr.2014.04.028
4. Naismith, J., Devine, T., Kohno, T. et al. Structures of the extracellular domain of the type I tumor necrosis factor receptor. Structure, 4 (11), 1251-1262, (1996). https://doi.org/10.1016/S0969-2126(96)00134-7
5. Vanamee, É. and Faustman, D. Structural principles of tumor necrosis factor superfamily signaling. Science Signaling, 11 (511), eaao4910, (2018) https://doi.org/10.1126/scisignal.aao4910
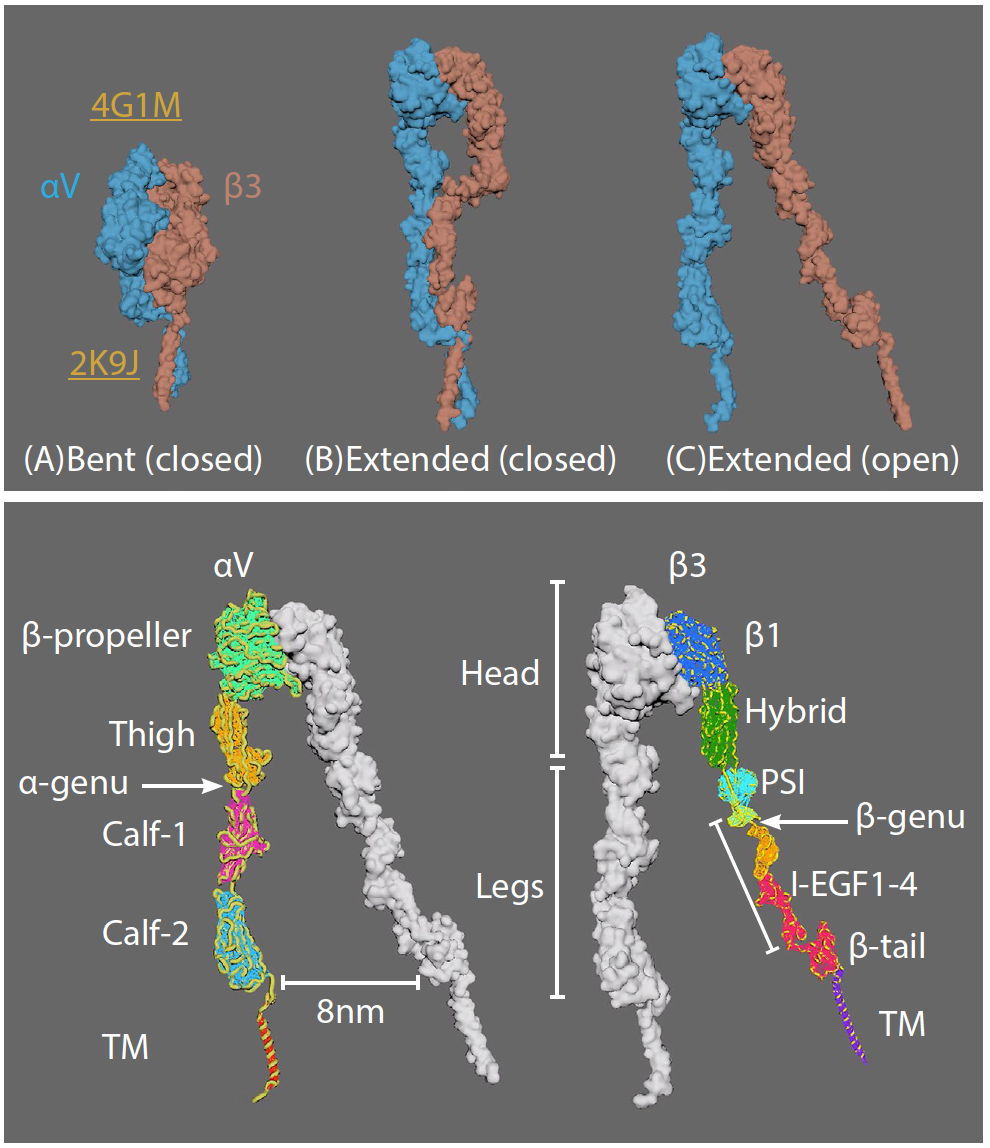
αVβ3 integrin
• Three major integrin conformational states: (A) bent with closed headpiece, (B) extended with a closed headpiece, and (C) extended with an open headpiece.
• Switchblade model: conformational change from bent to extended
• Extracellular domains: 4 in alpha; 8 in beta chains
• Bent conformation at knees/genua: between thigh and calf-1 in alpha5; between EGF1 and EGF2 in beta3
• Ligands vary (e.g. ECM proteins)
• Switchblade model: conformational change from bent to extended
• Extracellular domains: 4 in alpha; 8 in beta chains
• Bent conformation at knees/genua: between thigh and calf-1 in alpha5; between EGF1 and EGF2 in beta3
• Ligands vary (e.g. ECM proteins)
References:
1. Dong, X., Mi, L., Zhu, J. et al. αVβ3 Integrin Crystal Structures and Their Functional Implications. Biochemistry, 51 (44), 8814-8828 (2012). https://doi.org/10.1021/bi300734n
2. Lau, T., Kim, C., Ginsberg, M. et al. The structure of the integrin αIIbβ3 transmembrane complex explains integrin transmembrane signalling. The EMBO Journal, 28: 1351-1361, (2009). https://doi.org/10.1038/emboj.2009.63
3. Zhu, J., Luo B., Xiao, T. et al. Structure of a Complete Integrin Ectodomain in a Physiologic Resting State and Activation and Deactivation by Applied Forces. Molecular Cell, 32 (6), 849-861 (2008). https://doi.org/10.1016/j.molcel.2008.11.018
4. Chen, W., Lou, J., Hsin, J. et al. Molecular Dynamics Simulations of Forced Unbending of Integrin αVβ3. PLOS Computational Biology 7(2):e1001086, (2011). https://doi.org/10.1371/journal.pcbi.1001086
1. Dong, X., Mi, L., Zhu, J. et al. αVβ3 Integrin Crystal Structures and Their Functional Implications. Biochemistry, 51 (44), 8814-8828 (2012). https://doi.org/10.1021/bi300734n
2. Lau, T., Kim, C., Ginsberg, M. et al. The structure of the integrin αIIbβ3 transmembrane complex explains integrin transmembrane signalling. The EMBO Journal, 28: 1351-1361, (2009). https://doi.org/10.1038/emboj.2009.63
3. Zhu, J., Luo B., Xiao, T. et al. Structure of a Complete Integrin Ectodomain in a Physiologic Resting State and Activation and Deactivation by Applied Forces. Molecular Cell, 32 (6), 849-861 (2008). https://doi.org/10.1016/j.molcel.2008.11.018
4. Chen, W., Lou, J., Hsin, J. et al. Molecular Dynamics Simulations of Forced Unbending of Integrin αVβ3. PLOS Computational Biology 7(2):e1001086, (2011). https://doi.org/10.1371/journal.pcbi.1001086